|
AUGUSTOWSKO-SUWALSKIE TOWARZYSTWO NAUKOWE
Proszę chwilę zaczekać, ładuję stronę... |
|
AUGUSTOWSKO-SUWALSKIE TOWARZYSTWO NAUKOWE
Proszę chwilę zaczekać, ładuję stronę... |
|
Letni fitoplankton wybranych sucharów Wigierskiego Parku Narodowego
Zbiorniki dystroficzne są siedliskiem stwarzającym glonom planktonowym specyficzne warunki rozwoju (Eloranta 1986; Arvola i in. 1990; Hutorowicz 1993, 1999, 2001; Jansson i in. 1996; Luścińska, Soska 1998). Duża zawartość substancji humusowych przyczynia się do znacznego zakwaszenie i zacienienia wody. Ponadto substancje humusowe wiążą biogeny w trwałe chelaty, czyniąc je niedostępnymi dla fitoplanktonu (Górniak 1996, 1999a, b). Do tak trudnych warunków siedliskowych dostosowują się tylko nieliczne glony, przede wszystkim są to miksotrofy i heterotrofy (Eloranta 1986; Arvola i in. 1990; Jansson i in. 1996; Górniak i in. 1999a). Strefa występowania autotrofów ogranicza się głównie do powierzchniowej warstwy wody.
Teren i metody badań
W sierpniu 2002 roku badaniami objęto 10 sucharów z terenu WPN (tab. 1), czyli ponad połowę wszystkich jezior dystroficznych parku. Do badań algologicznych pobierano 300–500 cm3 wody ze śródjezierza. Próby w terenie utrwalano formaliną i płynem Utermöhl’a. Analizę jakościową i ilościową przeprowadzano w kamerach Fuchs-Rosenthal’a. Liczono co najmniej 100 osobników w próbie. Biomasę glonów określano metodą pomiaru objętości komórek.
Tabela 1. Lista badanych sucharów z głębokościami poboru prób
Badane suchary charakteryzują się znacznym zakwaszeniem i zacienieniem wody oraz niskim stężeniami biogenów. Głębsze zbiorniki posiadają wyraźną stratyfikację termiczną i tlenową.
Wyniki
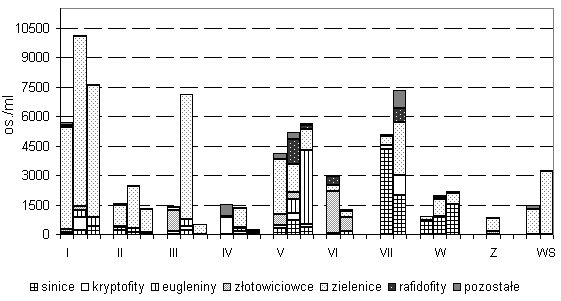
Letni fitoplankton sucharów reprezentowany był przez 9 grup glonów (rys. 2b). Liczebność fitoplanktonu (Lf) wykazywała bardzo duże zróżnicowanie – od 237 do 10 111 os./cm3. W większości sucharów Lf wzrastała wraz z głębokością, osiągając swoje maksimum na głębokości 1,5–2,5 m. O liczebności decydowały przede wszystkim drobne zielenice: Chlorella sp., Dictyosphaerium sp. div. w SI, SV, SVII, Monoraphidium contortum (Tur.) Kom.-Legn. w SI, SVII, Z, Chlamydomonas sp. w SII, SIII, SIV, Crucigenia tetrapedia (Kirchner) W. et G. S. West i Quadrigula koršikovii Komárek w SW. W sucharach SI, SII, SIII, Z i WS zielenice stanowiły ponad 70 proc. Lf. Złotowiciowce dominowały jedynie w warstwie powierzchniowej SIII (Dinobryon divergens Imhoff) i w SVI (Dinobryon bavaricum Imhoff), a sinice w SVII (Synechococcus sp.) i W (Me- rismopedia tennuissima Lemm./. Eugleniny, kryptofity i rafidofity stanowiły stały składnik fitoplanktonu badanych jezior, lecz miały mały udział w liczebności (rys. 1).
Rys. 1. Liczebność całkowita fitoplanktonu i udział
poszczególnych grup w sucharach WPN
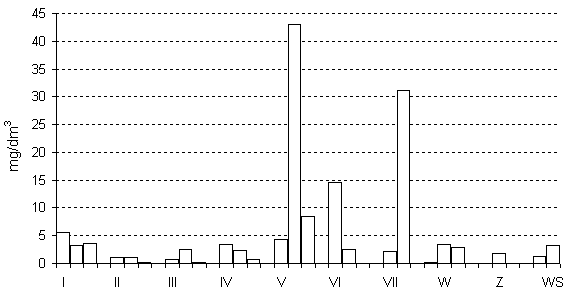
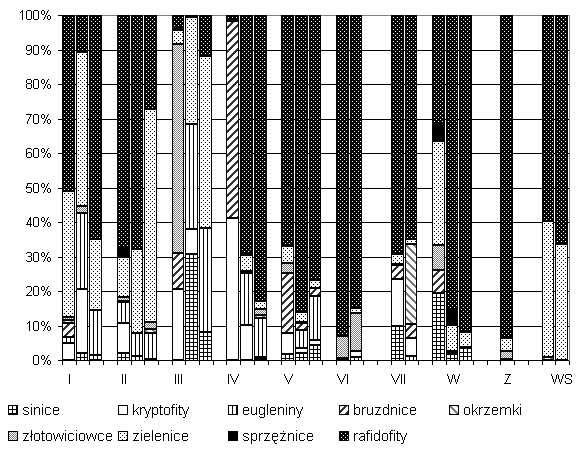
W SIII i SIV odnotowano największą zmienność składu taksonomicznego wraz z głębokością (rys.1, 2b). W SIII przy powierzchni największą Lf i biomasę osiągają złotowiciowce (Dinobryon divergens), a w SIV tobołki (Peridinium incospicuum Lemm.) nienotowane już w głębszych warstwach jezior. W większości badanych sucharów biomasa całkowita fitoplanktonu (Bf) nie przekraczała 5 mg/dm3, jedynie w SVI, SV i SVII wartości były zdecydowanie większe (maks. 43,1 mg/dm3) (rys. 2a). Średnia Bf w badanych sucharach wynosiła 4,16 mg/dm3. Największą biomasę prawie we wszystkich jeziorach osiągały rafidofity. Jedynie w SIII w warstwie przypowierzchniowej zdecydowanym dominantem były złotowiciowce, a w warstwach głębszych zielenice, eugleniny i sinice (rys. 2b).
Rys. 2 a, b. Biomasa całkowita fitoplanktonu (a) i
udział procentowy poszczególnych grup w biomasie fitoplanktonu (b) sucharów WPN
w sierpniu 2002 roku.
Istotnym składnikiem biomasy w większości sucharów były zielenice, kryptofity, eugleniny i bruzdnice (rys. 2b). Okrzemki notowano wprawdzie aż w siedmiu sucharach, ale jedynie w SVII (2 m) stanowiły one istotny składnik Bf (24 proc. Bf, 7,25 mg/dm3). Występujące w SVII okrzemki Pinnularia minor (Kütz.) Cleve i Tabellaria flocculosa (Roth) (Kütz.) nie były notowane w pozostałych sucharach. Najmniejsze liczebności najczęściej osiągały w zbiornikach sprzężnice (0–180 os./cm3) z rodzaju Euastrum (SVII), Cosmarium (SI, SII, SIV), Staurastrum (SII), a w Sucharze Wielkim Staurodesmus sp. i Arthrodesmus octocornis Ehr. Rafidofity reprezentowane były przez jeden tylko gatunek Gonyostomum semen (Ehr.) Diesing. Notowano go na wszystkich stanowiskach, przy czym największą liczebność i biomasę osiągnął w SV na głębokości 2 m (1250 os./cm3, 33 mg/dm3).
Dyskusja
W warunkach podwyższonej barwy i dużego zakwaszenia wody oraz niskich stężeń biogenów w zbiornikach dystroficznych (Górniak 1996; Górniak i in. 1999a, b, w tym tomie) notuje się słaby rozwój glonów. Największą liczebność i biomasę osiągają miksotrofy i heterotrofy (Eloranta 1986; Arvola i in. 1990; Jansson i in. 1996; Hutorowicz 1999, 2001). W jeziorach dystroficznych przepływ energii zapewnia zespół bakterii i miksotroficznych glonów z małym udziałem lub pominięciem autotrofów (Jansson i in. 1996). W sucharach WPN najliczniej reprezentowane były miksotroficzne wiciowce z rodzaju Gonyostomum, Dinobryon, Chlamydomonas, Euglena, Cryptomonas i Peridinium, które mogą korzystać z rozpuszczonej lub cząsteczkowej materii organicznej (Kawecka, Eloranta 1994). Podobny duży udział rafidofitów, bruzdnic, kryptofitów i zielenic w fitoplanktonie stwierdził wcześniej w 19 sucharach WPN Hutorowicz (1999). Złotowiciowiec Dinobryon bavaricum, osiągający dużą biomasę w SIII, notowany był przez Wesołowską (2003) w SI , W i WS. Mały udział desmidii w fitoplanktonie jezior dystroficznych potwierdzają wyniki badań Luścińskiej i Soski (1998) z Borów Tucholskich. Dla desmidii lepsze warunki rozwoju istnieją w pobliżu pła torfowcowego niż w śródjezierzu (Luścińska, Soska 1998). Duży udział okrzemek (SVII) i sinic (SVII, W) może wskazywać na powolną eutrofizację zbiorników (Eloranta 1986). Tego typu zbiorniki Eloranta (1986) określa mianem dyseutroficznych. Suchar VII i Suchar Wielki mają jednocześnie najniższe wskaźniki zaawansowania dystrofii na terenie WPN (Górniak, w tym tomie). Gonyostomum semen występujący na wszystkich stanowiskach tolerował bardzo szeroki zakres parametrów fizyczno-chemicznych wody (tab. 2). Występował zarówno w dobrze natlenionej, powierzchniowej warstwie wody, jak też w warstwach głębszych, zupełnie pozbawionych tlenu. W oparciu o maksymalną liczebność i biomasę można przyjąć, że Gonyostomum semen optymalne warunki rozwoju osiągnął na głębokości 2 m w SV (tab. 2). Również Hutorowicz (1993), w humusowym jeziorze Smolak, maksymalne liczebności Gonyostomum semen rzędu 5000–6000 os./cm3 stwierdził w metalimnionie.
Tabela 2. Zakres fizyczno-chemicznych parametrów wody badanych sucharów w czasie występowania Gonyostomum semen
Dużą tolerancję Gonyostomum semen w stosunku do panujących w środowisku wodnym warunków świetlnych, termicznych, tlenowych i pokarmowych potwierdzają również inne badania (Hutorowicz 1993, 1999, 2001). Tak zróżnicowane warunki bytowania tego taksonu świadczą o jego dużej plastyczności siedliskowej. Gatunek uznawany wcześniej za typowy dla dystrofii (Arvola i in. 1990; Hutorowicz 1993, 1999, 2001; Górniak i in. 1999a), obecnie coraz częściej notowany jest w jeziorach lobeliowych (Szeląg-Wasielewska, Gołdyn 1994), a także w zeutrofizowanych jeziorach dystroficznych i torfiankach (Hutorowicz 2001). Bardzo zmienne warunki środowiska wodnego na różnych głębokościach sucharów pociągają za sobą duże zmiany w składzie gatunkowym, liczebności i biomasie fitoplanktonu. Złotowiciowiec Dinobryon divergens i bruzdnica Peridinium incospicuum stwierdzane są tylko w warstwie powierzchniowej SIII i SIV. Wiciowce mają możliwość pionowego przemieszczania się w toni wodnej w celu kolonizacji dogodnych dla siebie siedlisk pod względem fizycznym i chemicznym. Miksotroficzne wiciowce mogą wybierać między zasobniejszym w biogeny, pozbawionym najczęściej światła metalimnionem i oświetlonym, ale uboższym w biogeny – epilimnionem. Tego typu wędrówki wykonuje Gonyostomum semen (za Hutorowicz 2001). Suchary III, IV i V o najniższym pH (4,5–5,4), charakteryzujące się największym udziałem miksotroficznych rafidofitów, kryptofitów, euglenin, złotowiciowców i bruzdnic (rys. 1, 2a, b), mają najwyższy wskaźnik zaawansowania dystrofii wśród wszystkich sucharów WPN.
Abstract
Summer phytoplankton of some dystrophic lakes of Wigry National Park Phytoplankton structure, abudance and biomass were studied in 10 small dystrophic lakes of Wigry National Park. Summer phytoplankton was dominated by mixotrophic flagellates from Raphidophyceae, Chlorophyceae, Cryptophyta, Pyrrophyta, Chrysophyceae and Euglenophyta. The most frequent species was Gonyostomum semen. It was recorded in all investigated lakes under diversified light, temperature, oxygen and nutrient conditions.
Literatura
Arvola L., Metsälä T.-R., Similä A., Rask M., 1990: Phyto- and zooplankton in relation to water pH and humic content in small lakes in Southern Finland. Verh. Internat. Verein. Limnol., 24:688–692. Eloranta P., 1986: Phytoplankton structure in different lake types in central Finland. Holarctic Ecology., 9:214–224. Górniak A., 1996: Substancje humusowe i ich rola w funkcjonowaniu ekosystemów słodkowodnych. Białystok, Diss. Univ. Varsoviensis, s. 448, 151. Górniak A., Grabowska M., Dobrzyń P., 1999a: Fitoplankton trzech jezior dystroficznych Wigierskiego Parku Narodowego. W: Zdanowski B., Kamiński M., Martyniak A. (red.): Funkcjonowanie i ochrona ekosystemów wodnych na obszarach chronionych. Olsztyn, Wydawnictwo IRS, s. 351–370. Górniak A., Jekatierynczuk-Rudczyk E., Dobrzyń P., 1999b: Hydrochemistry of Three Dystrophic Lakes in Northeastern Poland. Acta Hydrochim. et Hydrobiol., 27:12–18. Hutorowicz A., 1993: Gonyostomum semen (Raphidophyceae) in Lake Smolak in northern Poland. Fragm. Flor. Geobot., 38 (1):163–171. Hutorowicz A., Tunowski J., Zdanowski B., 1999: Dystroficzne jeziora Wigierskiego Parku Narodowego – struktura, funkcjonowanie i zagrożenia. W: Radwan S., Kornijów R. (red.): Problemy aktywnej ochrony ekosystemów wodnych i torfowiskowych w polskich parkach narodowych. Lublin, Wyd. UMCS, s. 219–231. Hutorowicz A., 2001: Fitoplankton humusowego jeziora Smolak. Idee Ekologiczne, 14, Ser. Zeszyty. Wyd. SORUS, Poznań, s. 130. Jansson, M., Blomqvist P., Jansson A., BergströmA. K., 1996: Nutrient limitation of bakterioplankton, autotrophic and mixotrophic phytoplankton, and heterotrophic nanoflagellates in Lake Örträsket. Limnol. Oceanogr., 41 (17):1552–1559. Kawecka B., Eloranta P., 1994: Zarys ekologii glonów wód słodkich i środowisk lądowych. Warszawa, PWN. Luścińska M., Soska R., 1998: Algal communities of humic acid sites: Biomass and diversity in lake water versus peat-mat water. Pol. J. Ecol., 46 (2):123–135. Szeląg-Wasielewska E., Gołdyn R., 1994: Zbiorowiska glonów w pelagialu jezior lobeliowych. W: Kraska M.: Jeziora lobeliowe. Charakterystyka, funkcjonowanie i ochrona. Cz. 1. Idee Ekologiczne 6, Ser. Szkice, 4:37–65. Wesołowska L., 2003: Chrysophyceae and Xanthophyceae of selected sukhars and a peat bog in village of Rosochaty Róg in the Wigry National Park. Acta Botanica Warmiae et Masuriae, 3:199–211.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||